1. 전사(Transcription)
: 전사는 두 줄의 DNA에서 외줄의 RNA가 합성되어 나오는 과정이다. 유전자발현의 첫 번째 과정이며, 발현에 있어서 가장 중요한 조절 과정이다. 전사 과정은 원핵세포와 진핵세포 간 핵막의 유무에 따른 근본적인 차이가 존재하는데 원핵세포는 핵막이 없기 때문에 세포질에 둥둥 떠 있는 DNA를 바로 전사시킬 수 있다.
하지만 진핵세포는 핵이 있기 때문에 이야기가 다를 수밖에 없다. 전사는 핵에서 일어나지만 필요한 리보솜들이 세포질에 있기 때문에 전사되어 만들어진 RNA가 세포질로 옮겨와야만 비로소 해석이 시작될 수 있기 때문이다. 따라서 진핵세포에서는 전사와 해석이 별개의 공간, 서로 다른 시간대에 일어난다는 차이가 있다.
전사 과정을 이해하기 위해 먼저 DNA의 두 줄을 용어적으로 구별할 필요가 있다. DNA가 복제될 때는 두 줄이 모두 주형으로 이용되었지만 전사에서는 한 줄만이 주형으로 사용되기 때문이다. 두 줄의 DNA가 서로 방향성이 다르기 때문에 국제 규약에 따르면 아무 표시가 없을 때 왼쪽 상단이 자연스럽게 5'이 된다.
RNA의 합성 방향도 5'→3'이므로 당연히 DNA의 두 가닥 중 상보적인 3'→5' 줄이 전사를 위한 주형이 된다. 따라서 3'→5' 줄을 주형줄(Template strand)이라 부르고 그 반대쪽 줄인 5'→3' 줄을 비주형줄(Non-template strand)이라고 부른다. 최근에는 실질적인 의미에 강조를 둔 용어를 사용하고 있다.
전사되어 나온 RNA에는 당연히 유전정보가 담겨 있다. 즉 DNA에 간직된 생명의 의미가 그대로 복사된 것이다. 이런 뜻에서 우리는 RNA를 센스 RNA(Sense RNA)라고 부른다. 그렇다면 당연히 RNA와 염기서열이 같은 5'→3' 줄, 즉 비주형줄이 센스 DNA(Sens DNA)가 될 것이고, 주형줄은 안티센스 DNA(Antisense DNA)가 되는 것이다. 따라서 RNA의 염기서열은 센스 DNA와 같고 다만 T(Tymine)가 U(Uracil)로 바뀌게 된다.


2. 진핵세포의 전사
: 진핵세포의 전사도 원핵세포에서와 크게 다르지는 않지만 전반적으로 훨씬 더 복잡하다. 일단 RNA 중합효소 Ⅰ,Ⅱ,Ⅲ의 세 종류이므로 유전자마다 이들이 구별하여 인식할 수 있도록 프로모터가 달라야 할 것이다. 즉 RNA 중합효소 Ⅱ는 mRNA의 유전자들을 인식하지만 Ⅰ번 효소가 인식하는 tRNA 유전자는 인식 못 한다는 것이다.
진핵세포에서 기본적인 프로모터는 TATA box이다. 하지만 유전자발현이 다양한 요소와 환경적 요인에 의해 변화하는 만큼 단순히 TATA box만으로 모두 조절될 수는 없다. 증폭자(Enhancer), 침묵자(Silencer) 등 새로운 염기서열들이 조절인자로서 밝혀져 있다.
| RNA 중합효소 종류 | 위치 | 합성하는 RNA |
| Ⅰ | 인 | rRNA(45S 전구체 RNA) |
| Ⅱ | 핵 | mRNA 전구체 대부분의 snRNA(U1,U2,U3 등) |
| Ⅲ | 핵 | tRNA, 5S rRNA, U6 snRNA 등 |
* TATA box
: 많은 진핵 세포의 유전자의 전사 개시점보다 위쪽으로 25~30 뉴클레오타이드에 위치하는 서열. 호그네스 박스라고도 한다. 전사 개시에 필요한 전사 인자 단백질 복합체와 아르엔에이(RNA) 중합 효소와 결합한다.
3. RNA 중합효소
: RNA도 고분자이기 때문에 중합효소(Polymerase)에 의해 만들어진다. RNA 중합효소는 다섯 개의 소단위(subunit)로 구성되어 있다. 이들은 각각 두 개의 α와 한 개의 β 그리고 β'과 σ이다. RNA 중합효소는 두 가지 형태를 취하고 있다. σ까지 붙어 α2 β β' σ의 온전한 구조를 취했을 때가 완전효소(Holo enzyme)로서 분자량은 480,000Da 정도이다. 그러나 σ 가 떨어져 나가서 α2 β β' 만 있게 되면 중심 효소(Core enzyme)가 된다. 이 두 가지의 서로 다른 형태는 전사 과정에서의 임무 역시 조금 다르다. 진핵세포의 RNA 중합효소는 조금 더 복잡하다. 종류도 많아서 최소한 세 가지의 RNA 중합효소가 이미 발견되어 있다. 이들은 각각 RNA 중합효소 Ⅰ,Ⅱ,Ⅲ로 이름 붙여졌는데 별개의 기능들을 가지고 있어 Ⅰ은 rRNA 합성을, Ⅱ는 mRNA 합성을 Ⅲ은 tRNA와 5s rRNA 그리고 U-RNA 합성을 담당하고 있다.
4. 전사 과정
: RNA의 전사 과정은 크게 세 단계로 구분할 수 있다. 개시 → 연장 → 종결
1) 개시
: 전사할 유전자의 시작 부분을 RNA 중합효소가 인식하는 것이다. 전사가 항상 일정한 곳에서 시작되고 또 끝나야만 온전한 단백질을 합성할 수 있기 때문이다. 유전자의 앞부분은 RNA 중합효소가 인식할 수 있도록 특별한 구조를 하고 있다. 이 부분을 전사를 촉진시킨다는 의미에서 프로모터(Promoter)라 부른다. 프로모터는 엄밀히 말해서 TATA 또는 이것이 약간 변형된 염기서열이다. 때로는 프로모터를 TATA box 또는 Pribnow box라고도 부른다.
전사가 시작되는 첫 번째 염기를 +1로 지칭한다. 그리고 프로모터는 이보다 상류 쪽, 즉 5' 쪽으로 10여 개 뉴클레오티드(-10) 정도 떨어져 있다. RNA 중합효소의 σ가 바로 이 프로모터를 인식할 수 있다. σ는 DNA를 스캐닝해 나가다가 프로모터를 발견하면 달라붙어서 여기가 바로 유전자의 시작임을 알려준다. 그러고는 자신의 의무를 다했기 때문에 완전효소로부터 떨어져 나가게 된다. 그러면 RNA 중합효소는 중심 효소만 남아 TATA box를 벌린 다음 +1지점으로부터 상보적 뉴클레오티드를 이어 나가기 시작한다.
프로모터의 염기서열이 TATA인 이유는 T와 A 사이에는 수소결합이 2개밖에 없으므로 RNA 중합효소가 비교적 쉽게 벌릴 수 있기 때문이다. 거의 모든 유전자에서 프로모터의 염기는 TATA이기 때문에 이를 공통 염기서열(Consensus sequence)이라 부른다.

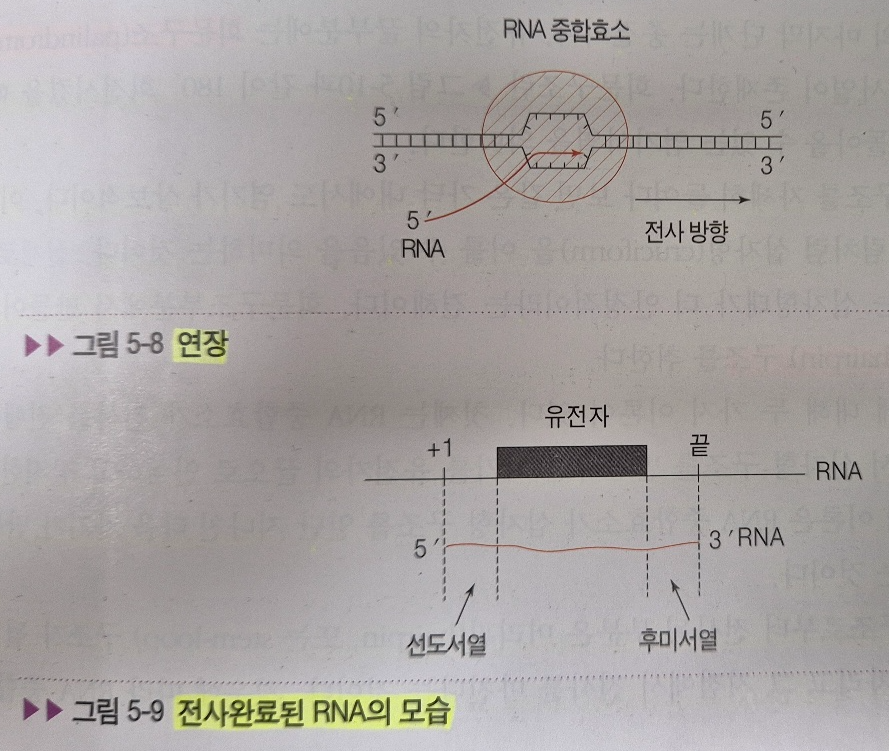
2) 연장
: TATA box를 인식하고, +1지점에 첫 번째 뉴클레오티드를 떨어뜨림으로써 본격적인 RNA 합성이 시작된다. 이 합성은 단순한 중합반응으로서 DNA 주형에 상보적인 염기를 계속 연장해 나가는 생화학 반응의 연속이다. 다만 주형이 A일 때 상보적 염기로서 T 대신 U를 집어넣는 것이 주의할 점이다. 연장 과정에서 3가지 중요한 부분이 있다.
첫 번째, 최종 RNA 산물에는 유전정보도 들어있지만 좌우에 여백이 있다는 것이다. 즉, 유전정보는 +1이 아닌, 그보다 훨씬 안쪽에 자리 잡고 있다는 것이다. 비록 +1부터 전사가 되더라도 실제 유전정보의 시작이 ATG 염기서열까지는 별로 특별한 의미가 없는 선도 서열(Leader sequence)이 있기 마련이다. 마찬가지로 유전정보가 끝난 3' 쪽에도 특별한 유전적 의미가 없는 후미 서열(Trailer sequence)이 붙어있다. 이 염기서열들은 마치 유전정보를 경호하듯이 좌우로 둘러싸고 있어 RNase 효소의 공격으로부터 보호해 주는 역할을 담당한다.
두 번째, RNA 중합효소는 DNA 주형을 풀어나가면서, 또 동시에 사슬을 연장시켜야 하는 대단히 바쁜 효소이다. 그렇기 때문에 교정 능력을 가지고 있지 않다. DNA라면 다음 세대까지 물려주어야 하는 중요한 유전정보이기 때문에 철저하게 교정하여 실수가 없도록 해야겠지만 RNA는 단백질을 합성하기 위한 중간 과정에 불과하고 또 똑같은 것이 수천 개씩 발현되고 있으므로 이 중 하나쯤 잘못 만들어질지라도 생명에는 별 지장이 없기 때문이다.
세 번째, RNA 중합효소는 헬리카아제와 달리 DNA 사슬을 한 번에 조금씩만 풀었다가 되감아 준다. 그럼으로써 막 합성이 끝난 RNA는 DNA 두 줄이 서로 원위치할 때 주형으로부터 떨어져 나올 수 있게 된다.
3) 종결
: 전사의 마지막 단계는 종결이다. 유전자의 끝부분에는 회문구조(Palindrome)라는 특별한 염기서열이 존재한다. 회문구조란 180˚ 회전시켰을 때 다시 원래 상태로 돌아올 수 있는 염기서열을 의미한다. 회문구조를 자세히 들여다보면 같은 가닥 내에서도 염기가 상보적이다. 이것은 회문구조가 그림처럼 십자형(Cruciform)을 이룰 수 있음을 의미하는 것이다. 회문구조에서 만들어지는 RNA는 머리핀(Hairpin)구조를 취한다.
종결에 대한 두 가지 이론이 있다. 첫째는 RNA 중합효소가 전사를 진행해 나가다가 DNA에서 십자형 구조를 발견하면 여기를 유전자의 끝으로 인식하고 정한다는 이론이다. 둘째 이론은 RNA 중합효소가 십자형 구조를 일단 지나친 다음 자기가 만든 RNA를 돌아본다는 것이다. 회문구조로부터 전사된 부분은 머리핀(Hairpin or Stem-loop) 구조가 될 것이므로 이를 알아차리고 그 지점에서 전사를 마친다는 것이다.
전사가 종결되면 새로 합성된 RNA와 중합효소 등이 모두 DNA 주형에서 떨어져 나간다. 중심 효소는 다시 σ와 결합하여 완전효소를 형성한 뒤 두 번째 전사에 다시 나서게 된다.

출처 : 임상 분자생물학(정문각)
오늘은 DNA를 복제하기 위해 가장 첫 번째 단계인 전사에 대해 알아보았습니다.
전사라는 단어를 처음 접하게 되면 떠오르는 이미지는 어떤 것들이 있으신가요?

제가 처음 떠올린 '전사'라는 단어에 대한 이미지는 전쟁 속에서 독보적인 용맹함을 보여주는 전사의 이미지가 떠올랐는데요. 하지만 우리는 지금 분자생물학 속에 DNA와 관련된 여러 가지 생명현상 중 하나인 '전사'를 이야기하고 있습니다.
사전 속 의미를 살펴보면 전사는 어떤 형태를 글로 옮겨적는 행위를 말하고 있습니다. 이를 분자생물학적으로 적용해 보면 내 몸속에서 생명현상을 발생시킬 수 있는 가장 작은 형태인 세포에서 시작을 해보자면 세포를 만들기 위해서 여러 가지 세포 소기관들이 필요하고 이 세포 소기관이 만들어지기 위해서 단백질들이 필요합니다. 우리가 음식물을 섭취하고 여러 가지 생리작용을 통해 몸속에서 떠돌고 있는 단백질들을 그 기능과 목적에 맞게 조합을 해서 세포소기관들도 만들고 이런 소기관들을 조합해 세포를 만들어내야 합니다.

그렇다면 단백질들을 어떻게 기능에 맞게 조립할 수 있는지에 대해서는 저번 포스팅에서 설명해 드린 것처럼 목적으로 하는 설계대로 단백질들 조합하기 위한 설명서가 필요한데 이 설명서 같은 역할을 하는 것이 DNA입니다. 설명서인 DNA를 토대로 단백질을 만들어야 하는데 이 과정을 다음 포스팅에서 다루기로 하고 지금은 그 전 단계가 필요합니다. 설명서를 보고 만들어야 하는데 이 설명서는 절대로 잃어버려서는 안 됩니다. 그렇기 때문에 핵막 안에 안전하게 보존되어 있는 겁니다. 원본을 안전하게 보관하면서 결과물을 만들어내 우리는 사본을 발행하게 됩니다.
유명한 화가들이 그린 걸작들을 직접 보면 좋겠지만 그럴 수 없을 때 우리는 복사본이라도 접해보고 감상을 할 수 있는 것처럼 DNA도 마찬 가진 거죠. 원본은 보존하되 사본으로 목적을 달성할 수 있게 도와주는 것 그것이 전사 과정이라고 생각하면 좋을 것 같습니다.
단백질을 만들고 더 나아가 하나의 세포가 두 개로 분열하고 또 분열하며 개체수를 늘려가면서 생장하며 세포가 조직이 돼서 조직이 장기가 되고 장기가 기관이 되고 기관이 모여 하나의 생명체로 성장하는 이 과정에서 세포가 분열할 때 원본인 DNA가 핵막을 뚫고 나와 복제 과정을 거친다면 원본이 소실되어 똑같은 세포가 만들어지는 게 아니라 분열할 때마다 다른 세포가 만들어질 수도 있고, 그렇게 되면 제대로 된 하나의 생명체가 만들어지기 어렵겠죠?
그래서 진핵세포, 즉 핵막을 가지고 있는 생명체들은 DNA를 온전하게 보전하고 이 DNA의 복제본인 단일 가닥인 RNA를 만들어 핵막 밖으로 내보내 이 복사본을 가지고 번역(Translation) 과정을 거쳐 단백질들이 만들어지게 설계되어 있습니다.
이런 거 보면 주님께서 지혜롭게 인간을 설계하신 것 같습니다.
Transcription(전사)이라고 명명한 이 과정에 대해서 이해하는 데 오늘 포스팅이 도움이 되셨을까요?
제가 포스팅을 하는 목적은 어렵게 느껴지는 이론적인 과정을 좀 더 이해하기 쉬운 예를 들며 설명해 보고자 글을 풀어가는데 이해하시는 데 도움이 되셨으면 좋겠습니다.
다음은 Translation 과정에 대한 포스팅으로 찾아뵙도록 하겠습니다.
'분자생물학' 카테고리의 다른 글
| 유전자 조작(Gene Manipulation) 첫 단추 - 핵산 추출 (0) | 2024.02.22 |
|---|---|
| 유전자 발현의 조절 (Regulation of Gene Expression)과 돌연변이(Mutation) (0) | 2024.02.13 |
| 유전암호와 해석 (Genetic Code & Translation) (0) | 2024.02.05 |
| DNA 복제의 중요한 요소와 과정 (0) | 2024.01.26 |
| 분자생물학과 핵산에 대해 돌아보기 (0) | 2024.01.19 |



